
Our paper Rod-cone crossover connectome of mammalian bipolar cells has been republished in a special issue of The Journal Of Comparative Neurology, Retinal Special Issue I: Mammals.
Our paper Rod-cone crossover connectome of mammalian bipolar cells has been republished in a special issue of The Journal Of Comparative Neurology, Retinal Special Issue I: Mammals.

The Marclab for Connectomics is off to RD2018 and ISER 2018 in Killarney, Ireland and Belfast, Northern Ireland. I’ll be organizing sessions on retinal degeneration, and I’m tremendously proud of the work Dr. Crystal Sigulinsky will be presenting from her work on gap junctional connectivity in retinal degenerations and the work Dr. Rebecca Pfeiffer (@BeccaPfeiffer19) will be presenting on her work on the retinal pathoconnectome in two talks on bipolar cells and Müller cells.
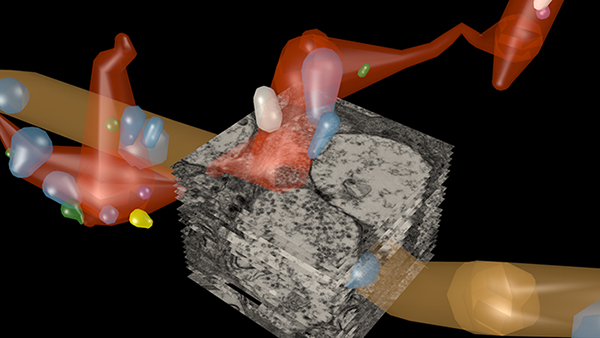
This abstract was presented at the 2014 FASEB Summer Research Conference in Saxtons River, Vermont by J. Scott Lauritzen, Crystal L. Sigulinsky, Noah T. Nelson, Nathan R. Sherbotie, Danny P. Emrich, Rebecca L. Pfeiffer, Jefferson R. Brown, John V. Hoang, Joshua M. Dudleston, Carl B. Watt, Kevin Rapp, Marguerite V. Shaw, Jia-Hui Yang, James R. Anderson, Bryan W. Jones and Robert E. Marc.
Purpose: Functional mapping in tiger salamander shows that bipolar cell (BC) projective fields far exceed their axonal fields, and directly implicates wide-field GABAergic amacrine cells (wf γACs) and gap junctions (Asari & Meister, 2014). Strikingly, single BCs exert differential effects on functionally distinct ganglion cells (GCs), likely achieved by privatized amacrine cell (AC) presynaptic inhibition to specific BC-GC synaptic pairs (Asari & Meister, 2012). To address whether BC projective fields in the mammal are equally broad, wf γAC- and gap junction-dependent, and GC type unselective, we reconstructed all electrical and chemical synaptic partners of a single ON cone BC in the inner plexiform layer of the rabbit retina, and searched BC-GC synaptic pairs for differential synaptic inhibition.
Methods: Cells in retinal connectome 1 (RC1) were annotated with Viking viewer, and explored via connectivity visualizations and 3D rendering (Anderson et al., 2011). Small molecule signals embedded in RC1, e.g. GABA, glycine, and L-glutamate, combined with morphological reconstruction and connectivity analysis allow robust cell classification. We used the MacNeil et al. (2004) rabbit BC classification scheme.
Results: CBb5w 593 is one of 20 ON cone BCs of this class in RC1. This CBb5w is presynaptic to 17 distinct GCs and 262 AC processes, and postsynaptic to 228 AC processes. The majority of these ACs are wf γACs. We estimate this BC forms synapses with 50 unique ACs. Asari & Meister (2014) found that single bipolar cell projective fields range up to 1 mm, far beyond a BC axonal field, and differentially drive multiple classes of GC. We discovered BC-BC within- and cross-class coupling and lateral inhibition that construct sign-conserving and sign-inverting projective fields to many distinct ganglion cell classes across the entire 0.25 mm diameter of RC1, much greater than a 60 µm BC axonal field. Cross-class projections access a broader set of GCs than expected from in-class projections alone. The BC-BC coupling is independent of BC-AII AC coupling. 94% of the CBb5w 593 BC-GC synaptic pairs receive feedback inhibition within the varicosity of the ribbon, but the number of feedback synapses is highly variable (coefficient of variation = 0.81). 35% of the BC-GC pairs receive feedforward inhibition within 2 microns of the postsynaptic density.
Conclusions: Mammalian BCs use novel cross-class topologies to distribute signals to a wide range of GCs and establish projective fields similar to those discovered in non-mammalian species. BC-BC within- and cross-class coupling and lateral inhibition via wf γACs establish sign-conserving and sign-inverting projective fields, respectively, up to 1 mm diameters. BC-GC synaptic pairs overwhelmingly employ feedback vs. feedforward inhibition to modulate signaling, and the numbers of feedback synapses are highly variable across these pairs, accounting for privatized and differential GC responses to the same BC drive.
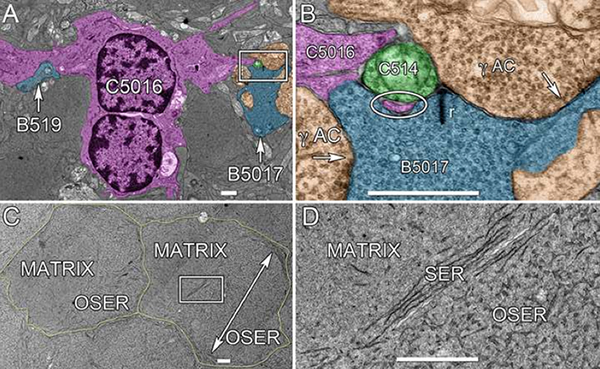
We have a new publication, Retinal connectomics: Toward complete, accurate networks in Progress in Retinal and Eye Research. Authors are: Robert E. Marc, Bryan W. Jones, Carl B. Watt, Crystal Sigulinsky, James R. Anderson and J. Scott Lauritzen.
Abstract:
Connectomics is a strategy for mapping complex neural networks based on high-speed automated electron optical imaging, computational assembly of neural data volumes, web-based navigational tools to explore 1012-1015 byte (terabyte to petabyte) image volumes, and annotation and markup tools to convert images into rich networks with cellular metadata. These collections of network data and associated metadata, analyzed using tools from graph theory and classification theory, can be merged with classical systems theory, giving a more completely parameterized view of how biologic information processing systems are implemented in retina and brain. Networks have two separable features: topology and connection attributes. The first findings from connectomics strongly validate the idea that the topologies complete retinal networks are far more complex than the simple schematics that emerged from classical anatomy. In particular, connectomics has permitted an aggressive refactoring of the retinal inner plexiform layer, demonstrating that network function cannot be simply inferred from stratification; exposing the complex geometric rules for inserting different cells into a shared network; revealing unexpected bidirectional signaling pathways between mammalian rod and cone systems; documenting selective feedforward systems, novel candidate signaling architectures, new coupling motifs, and the highly complex architecture of the mammalian AII amacrine cell. This is but the beginning, as the underlying principles of connectomics are readily transferrable to non-neural cell complexes and provide new contexts for assessing intercellular communication.